Resumindo este tópico do Evolution News… >>>> “O que é realmente a Teoria do Design Inteligente?”
Quem é o designer?
O que faz o designer?
Como é que ele faz?
Onde ele faz?
Quando ele faz?

Muitos críticos do ID promovem versões falsas, espantalhos da TDI:
“O design inteligente afirma que a vida é tão complexa, que não poderia ter evoluído, portanto, ela foi projetada por uma inteligência sobrenatural.”
Bom,
Parte A… O que o design inteligente não é.
1. ID não é somente um argumento negativo contra evolução.
ID não é apenas mero argumento contra evolução, ID oferece um forte argumento positivo, baseando-se em encontrar na natureza o tipo de informação e complexidade que vem somente de inteligência (baseando-se em nossa experiência).
2. ID não é uma teoria sobre o designer ou sobre o sobrenatural.
É um dos erros dos críticos, sugerir que a teoria está focada em estudar o designer; mais especificamente forças sobrenaturais ou uma divindade. Quando o ID estuda objetos naturais para determinar se eles carregam uma assinatura informativa indicando uma causa inteligente.
ID não se propõe a identificar a natureza ou a identidade dessa causa.
Como William Dembski explica:
“O design inteligente é a ciência que estuda os sinais de inteligência. Note que um sinal não é a coisa significada ….. Como um programa de pesquisa científica, design inteligente investiga os efeitos da inteligência, não a inteligência como tal.” [1]
Michael Behe explica:
“Muitas pessoas (inclusive eu) vão atribuir o projeto a Deus – com base, em parte, em outros, julgamentos não científicos que fizeram – eu não afirmo que a evidência bioquímica leva inevitavelmente a uma conclusão sobre quem é o designer . Na verdade, eu disse diretamente que, de um ponto de vista científico, a questão permanece em aberto. … A evidência bioquímica indica fortemente design, mas não mostra aonde o designer estava.” [2]
3. ID não é uma teoria de tudo.
ID é uma teoria científica de detecção de design, e isso é tudo.
ID não é uma teoria em pleno desenvolvimento, sobre tudo.Quem esperar ou exigir que o ID explique tudo sobre a história da vida e do cosmos, vai se decepcionar.
Se você quer saber se algo foi projetado ou não, tudo bem, volte-se para o ID.
Parte B... O que é o design inteligente.
1. ID utiliza argumento positivo baseado em encontrar elevados níveis de informação complexa e especificada.
A teoria do design inteligente começa com observações de como agentes inteligentes agem quando eles projetam coisas. A inteligência humana proporciona um grande conjunto de dados empíricos para estudar os produtos da ação de agentes inteligentes. Este conjunto de dados, baseado em observação atual estabelece relações de causa e efeito entre a ação inteligente e certos tipos de informação.
William Dembski observa que “[o] princípio característico da agência inteligente é contingência dirigida, ou o que chamamos de escolha.” [3] Dembski chama o ID de “uma teoria da informação”, onde “a informação torna-se um indicador confiável de design, bem como um objeto adequado para a investigação científica.“ [4] A relação de causa e efeito pode ser estabelecida entre mente e informações. Como o teórico da informação Henry Quastler observou, a “criação de novas informações é habitualmente associada à atividade consciente.” [5]
2. O projeto inteligente é uma ciência histórica que é metodologicamente equivalente ao neo darwinismo.
Como já vimos, o design inteligente é essencialmente uma ciência histórica, o que significa que estuda as causas atuais e, em seguida, as aplica ao registro histórico para inferir a melhor explicação para a origem dos fenômenos naturais. O design inteligente usa o raciocínio uniformista com base no princípio de que “o presente é a chave para o passado.”
Darwinistas usam este método para mutações e seleção. Afim de reconhecer capacidades causais e efeitos no mundo atual.Em seguida, tentam explicar o registro histórico em termos dessas causas, por exemplo buscando a reconhecer os efeitos conhecidos da mutação e seleção no registro histórico.
O design inteligente aplica esse mesmo método, estudando causas como a inteligência, a fim de reconhecer as suas capacidades causais e efeitos no mundo atual. Os teóricos do DI estão interessados em compreender os poderes de informação-generativa de agentes inteligentes. Os teóricos do DI, em seguida, tentam explicar o registro histórico, incluindo apelos para essa causa, procurando reconhecer os efeitos conhecidos de design inteligente (por exemplo, alta CSI) no registro histórico.
Então, se nós apelarmos para causas materialistas como mutação e seleção, ou causas não materiais, como o design inteligente, estamos usando o mesmo raciocínio uniformista básico e métodos científicos que são bem aceitos em ciências históricas. ID e neo-darwinismo são, portanto, metodologicamente equivalentes, o que significa que ambos são ou ciência, ou ambos não são ciência. No entanto, podemos saber que ID é ciência, porque ele usa o método científico.
3. O design inteligente usa o método científico.
ID usa o método científico para fazer suas reivindicações. Este método é comumente descrito como um processo de quatro etapas de: observações, hipóteses, experimentos e conclusão. Agora vou ilustrar isto referindo-se a quatro áreas científicas: bioquímica, paleontologia, sistemática e genética.
° ID e Bioquímica:
Observação: Os agentes inteligentes resolvem problemas complexos, atuando com um objetivo final em mente, produzindo altos níveis de CSI. Em nossa experiência, os sistemas com grandes quantidades de complexidade específica – como códigos e linguagens – invariavelmente são originários de uma fonte inteligente. Da mesma forma, em nossa experiência, a inteligência é a única causa conhecida de máquinas irredutivelmente complexos. [6]
Hipótese (Previsão): estruturas naturais que contêm muitas peças dispostas em intrincados padrões (incluindo a complexidade irredutível) que realizam uma função específica – indicando altos níveis de CSI.
Experiência: investigações experimentais de DNA indicam que ele é composto de um código baseado em linguagem rica em CSI. Os biólogos realizaram testes de sensibilidade mutacionais em proteínas e determinaram que as suas sequências de aminoácidos são altamente especificadas. [7] Além disso, experimentos genéticos inesperados e outros estudos têm mostrado que algumas máquinas moleculares, como o flagelo, são irredutivelmente complexas. [8]
Conclusão: Os altos níveis de CSI – incluindo a complexidade irredutível – em sistemas bioquímicos são melhor explicadas pela ação de um agente inteligente.
° ID e Paleontologia:
Observação: Os agentes inteligentes infundem rapidamente grandes quantidades de informação em sistemas. Como quatro teóricos do DI escreveram: “design inteligente fornece uma explicação causal suficiente para a origem de grandes quantidades de informação … o design inteligente de um projeto muitas vezes precede a montagem de peças de acordo com um projeto ou plano de projeto preconcebido.” [9]
Hipótese (Previsão): Formas que contêm grandes quantidades de novas informações aparecem no registro fóssil de repente e sem precursores semelhantes.
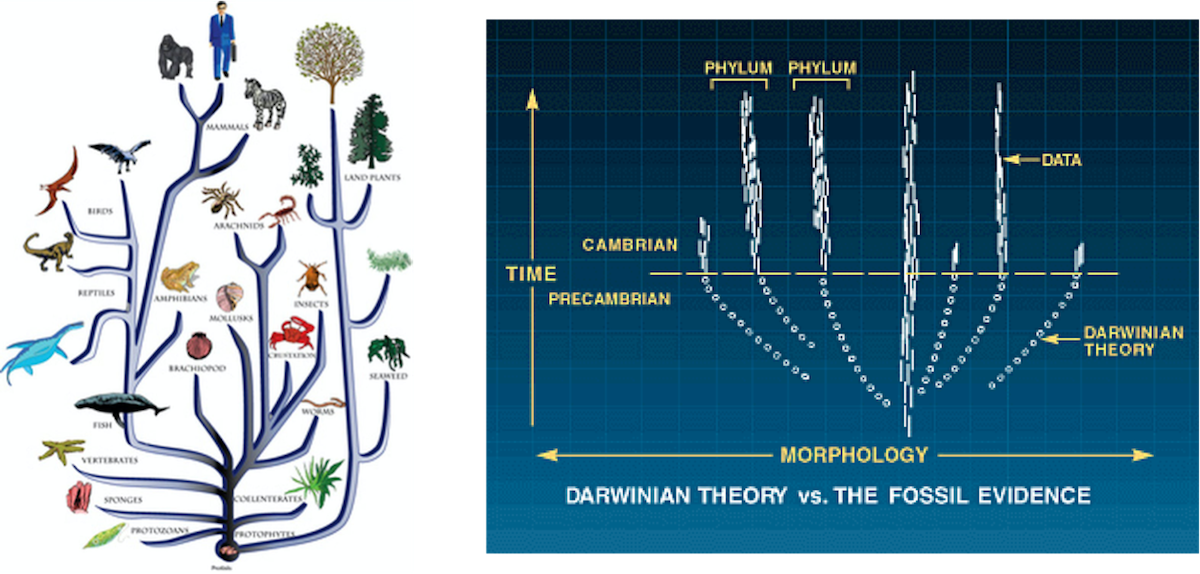
Experiência: Estudos sobre o registro fóssil mostram que as espécies geralmente aparecem de forma abrupta, sem precursores semelhantes. [10] A explosão cambriana é um excelente exemplo, embora existam outros exemplos de explosões na história da vida. Grandes quantidades de informações complexas e especificadas tiveram que surgir rapidamente para explicar o aparecimento abrupto dessas formas.[11]
Conclusão: O aparecimento abrupto de novos planos corporais totalmente formados no registro fóssil é melhor explicado por design inteligente.
° ID e Sistemática:
Observação: Os agentes inteligentes, muitas vezes reutilizam componentes funcionais em diferentes projetos. Como Paul Nelson e Jonathan Wells explicam: “. Uma causa inteligente pode reutilizar ou reimplantar o mesmo módulo em sistemas diferentes … [e] gerar padrões idênticos de forma independente” [12]
Hipótese (Previsão): Os genes e outras partes funcionais, normalmente, serão reutilizados em diferentes organismos. [13]
Experiência: Estudos de anatomia comparativa e genética descobriram peças semelhantes comumente existentes em organismos muito diferentes. Exemplos de “evolução extrema convergente” mostram reutilização de genes funcionais e estruturas de um modo não previsto pela ancestralidade comum.[14]
Conclusão: A re-utilização de partes altamente complexas e semelhantes, em organismos amplamente diferentes do padrão de árvore (arvore da vida) é melhor explicado através da ação de um agente inteligente.
° ID e Genética:
Observação: Os agentes inteligentes constroem estruturas com finalidade e função. Como William Dembski argumenta: “Considere o termo ‘DNA lixo’. … Em uma visão evolucionista esperamos uma grande quantidade de ADN inútil. Se, por outro lado, os organismos foram concebidos, esperamos que o ADN, tanto quanto possível,venha exibir função “. [15]
Hipótese (Previsão): Muito do chamado “DNA lixo” vai revelar que desempenha funções valiosas.
Experiência: Numerosos estudos têm descoberto funções no “DNA lixo”. Exemplos incluem funções para pseudogenes, íntrons e DNA repetitivo. [16]
Conclusão: A descoberta da função para vários tipos de “DNA lixo” foi prevista com sucesso pelo design inteligente.
Desta forma, podemos verificar que o design inteligente é uma teoria científica de boa-fé que usa o método científico para fazer suas reivindicações em vários campos científicos.
Referências usadas neste artigo:
[1.] William Dembski, The Design Revolution (InterVarsity Press, 2004), p. 33.
[2.] Michael Behe, “Philosophical Objections to Intelligent Design: Response to Critics,” (July 31, 2000) at
[3] William A. Dembski, The Design Inference: Eliminating Chance through Small Probabilities (Cambridge University Press 1998), p. 62.
[4] William A. Dembski, “Intelligent Design as a Theory of Information,” in Intelligent Design Creationism and Its Critics: Philosophical, Theological, and Scientific Perspectives (Robert T. Pennock ed., MIT Press 2001), p. 553.
[5] Henry Quastler, The emergence of biological organization, (Yale University Press, 1964), p. 16.
[6] Scott A. Minnich and Stephen C. Meyer, “Genetic analysis of coordinate flagellar and type III regulatory circuits in pathogenic bacteria,” Proceedings of the Second International Conference on Design & Nature, Rhodes Greece, edited by M.W. Collins and C.A. Brebbia (WIT Press, 2004).
[7] Douglas D. Axe, “Extreme Functional Sensitivity to Conservative Amino Acid Changes on Enzyme Exteriors,” Journal of Molecular Biology, Vol. 301:585-595 (2000); Douglas D. Axe, “Estimating the Prevalence of Protein Sequences Adopting Functional Enzyme Folds,” Journal of Molecular Biology, 1-21 (2004); Ann K Gauger, Stephanie Ebnet, Pamela F Fahey, Ralph Seelke, “Reductive Evolution Can Prevent Populations from Taking Simple Adaptive Paths to High Fitness,” BIO-Complexity, Vol. 2010; Ann K. Gauger and Douglas D. Axe, “The Evolutionary Accessibility of New Enzyme Functions: A Case Study from the Biotin Pathway,” BIO-Complexity, Vol. 2011(1) (2011).
[8.] See Kitzmiller Transcript of Testimony of Scott Minnich pp. 99-108, November 3, 2005; Robert M. Macnab, “Flagella,” in Escherichia Coli and Salmonella Typhimurium: Cellular and Molecular Biology Vol. 1, eds. Frederick C. Neidhardt, John L. Ingraham, K. Brooks Low, Boris Magasanik, Moselio Schaechter, and H. Edwin Umbarger (Washington D.C.: American Society for Microbiology, 1987), pp. 73-74.
[9.] Stephen C. Meyer, Marcus Ross, Paul Nelson, and Paul Chien, “The Cambrian Explosion: Biology’s Big Bang,” in Darwinism, Design, and Public Education, eds. John A. Campbell and Stephen C. Meyer (East Lansing, MI: Michigan State University Press, 2003), pp. 367, 386.
[10.] See Meyer, Ross, Nelson, and Chien, “The Cambrian Explosion: Biology’s Big Bang;” Wolf-Ekkehard Lönnig, “Dynamic genomes, morphological stasis, and the origin of irreducible complexity,” Dynamical Genetics, eds. Valerio Parisi, Valeria De Fonzo, and Filippo Aluffi-Pentini (Kerala, India, Research Signpost, 2004), 101-119; A.C. McIntosh, “Evidence of Design in Bird Feathers and Avian Respiration,” International Journal of Design & Nature and Ecodynamics, Vol. 4: 154-169 (2009).
[11.] Meyer, “The origin of biological information and the higher taxonomic categories.”
[12.] Paul Nelson and Jonathan Wells, “Homology in Biology,” in Darwinism, Design, and Public Education, eds. John Angus Campbell and Stephen C. Meyer (East Lansing: Michigan State University Press, 2003), p. 316.
[13.] In this case of systematics, neo-Darwinism might make some of the same predictions. Is this a problem for the positive case for design? Not at all. The fact that another theory can explain some data does not negate ID’s ability to successfully predict what we should find in nature. After all, part of making a “positive case” means that the arguments for design stand on their own and do not depend on refuting other theories. Moreover, there are many cases of supposed extreme “convergent evolution” that are better explained by common design. Additionally, regarding the predictions from biochemistry), paleontology, and genetics, neo-Darwinism has made different predictions from ID. In any case, in this example ID makes a slightly different prediction in that it does not predict that re-usage of parts must necessarily occur in a nested hierarchical pattern–a prediction which is in fact confirmed. See chapters 5-6 in Stephen C. Meyer, Darwin’s Doubt: The Explosive Origin of Animal Life and the Case for Intelligent Design (HarperOne, 2013).
[14.] John A. Davison, “A Prescribed Evolutionary Hypothesis,” Rivista di Biologia / Biology Forum, Vol. 98 (2005): 155-166; Nelson and Wells, “Homology in Biology;” Lönnig, “Dynamic genomes, morphological stasis, and the origin of irreducible complexity;” Michael Sherman, “Universal Genome in the Origin of Metazoa: Thoughts About Evolution,” Cell Cycle, 6: 1873-1877 (August 1, 2007).
[15.] William A. Dembski, “Science and Design,” First Things, Vol. 86 (October, 1998).
[16.] See Jonathan Wells, The Myth of Junk DNA (Discovery Institute Press, 2011); Richard Sternberg, “On the Roles of Repetitive DNA Elements in the Context of a Unified Genomic-Epigenetic System,” Annals of the NY Academy of Science, Vol. 981: 154-188 (2002); James A. Shapiro, and Richard Sternberg, “Why repetitive DNA is essential to genome function,” Biological Reviews of the Cambridge Philosophical Society, Vol. 80: 227-250 (2005); A.C. McIntosh, “Information and Entropy–Top-Down or Bottom-Up Development in Living Systems?,” International Journal of Design & Nature and Ecodynamics, Vol. 4: 351-385 (2009); The ENCODE Project Consortium, “An integrated encyclopedia of DNA elements in the human genome,” Nature, Vol. 489: 57-74 (September 6, 2012).